Was ist alternatives spleißen in der Genetik?
Alternatives Spleißen (engl. alternative splicing) ist ein genetischer Mechanismus bei Eukaryoten (Lebewesen mit Zellkern) mit Hilfe dessen aus einem einzigen Gen mehrere verschiedene Proteine hergestellt werden können.
Es gibt verschiedene Varianten des alternativen Spleißen:

Es ist der Hauptgrund, warum der Mensch mit nur rund 20.000 Genen weit über 100.000 verschiedene Proteine bilden kann.
Das Thema alternatives Spleißen wird in der Schule in der Oberstufe des Gymnasiums gelehrt. Je nach Bundesland und Schulsystem (G8 oder G9) entspricht dies meistens der Klassenstufe 11 oder 12 (selten auch bereits in der Klasse 10 am Ende der Einführungsphase). Das Problem, das von den Firmen ignoriert wurde, ist Schulstoff!!!
Alternative Spleißvarianten des Spike Proteins
Die meisten Menschen gehen davon aus, dass nach einer modRNA oder Adenoviren Injektion gegen COVID auch nur ein Spike-Protein produziert wird.
Dass dem nicht so ist und hybride Varianten entstehen, das habe ich bereits in diesem Artikel behandelt:
Das heißt, selbst wenn nur “normale”, vollständige Spike Monomere produziert würden, hat man durch die Rekombination der Untereinheiten bei Kreuzimpfungen oder jährlichen Updates bei bestehender Spike-Produktion eine unübersichtliche Anzahl an Immunogenen.
Johnson&Johnsen selbst haben gemerkt, dass es bei ihrem Genkonstrukt in eukaryotischen Zellen zu unterschiedlich gespleißten Spike-Varianten kommt.
Man fand vier verschiedene Splice-Events.

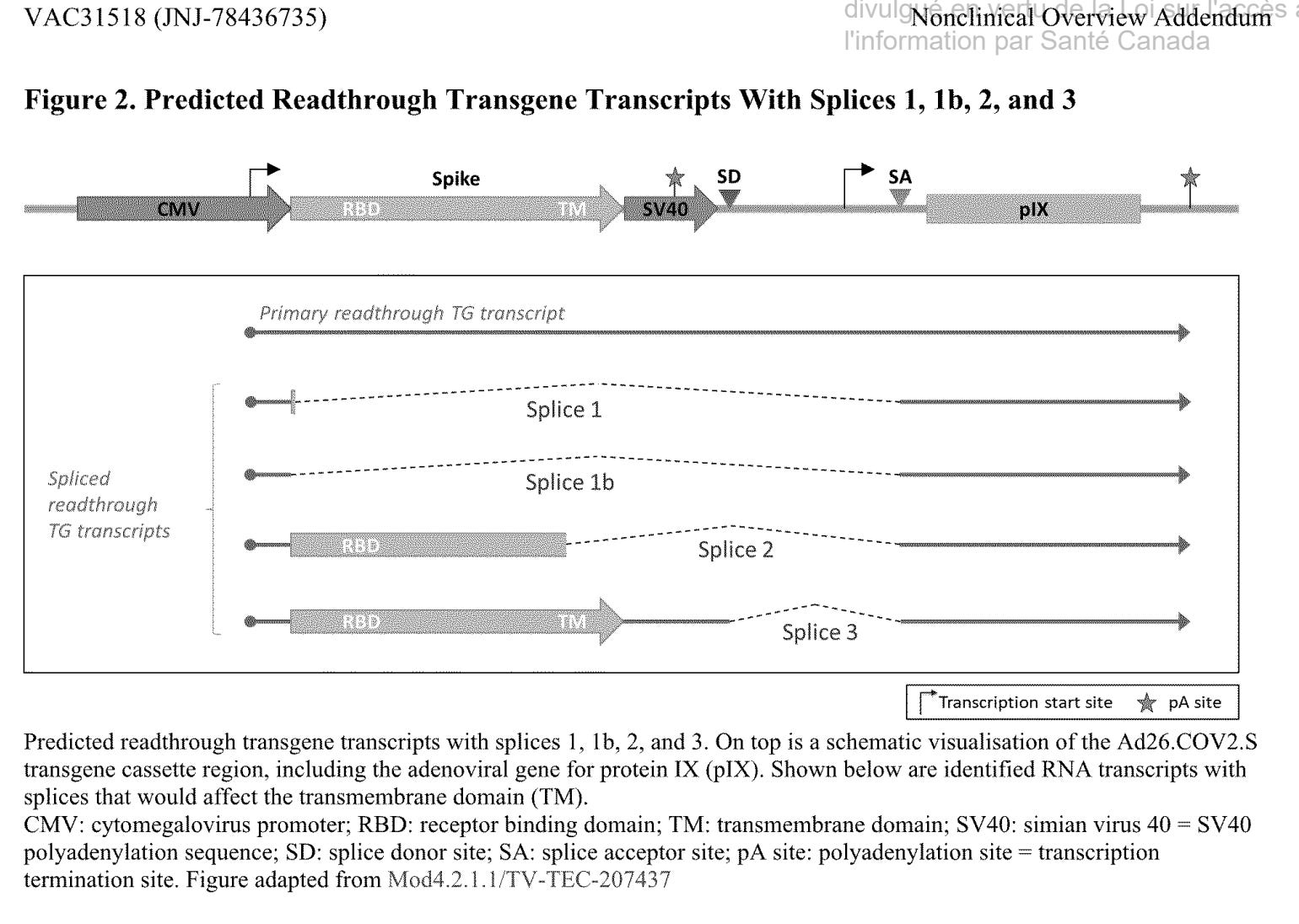
Diese gespleißten Proteine sind natürlich auch Antigene und/oder Immunogene, welche eine Immunreaktion hervorrufen bei gleichzeitiger hoher Homologie zu körpereigenen Proteinen. Das nennt man molekulare Mimikry. Die Details habe ich vor langer Zeit erklärt.
Es gibt eine Publikation aus dem Jahr 2022 die sich das Problem des alternativen Spleißens näher angesehen hat, die leider kaum jemand zur Kenntnis genommen hat.
Kowarz E, Krutzke L, Külp M, Streb P, Larghero P, Reis J, Bracharz S, Engler T, Kochanek S, Marschalek R. Vaccine-induced COVID-19 mimicry syndrome. Elife. 2022 Jan 27;11:e74974. doi: 10.7554/eLife.74974. PMID: 35084333; PMCID: PMC8846585. https://pmc.ncbi.nlm.nih.gov/articles/PMC8846585/
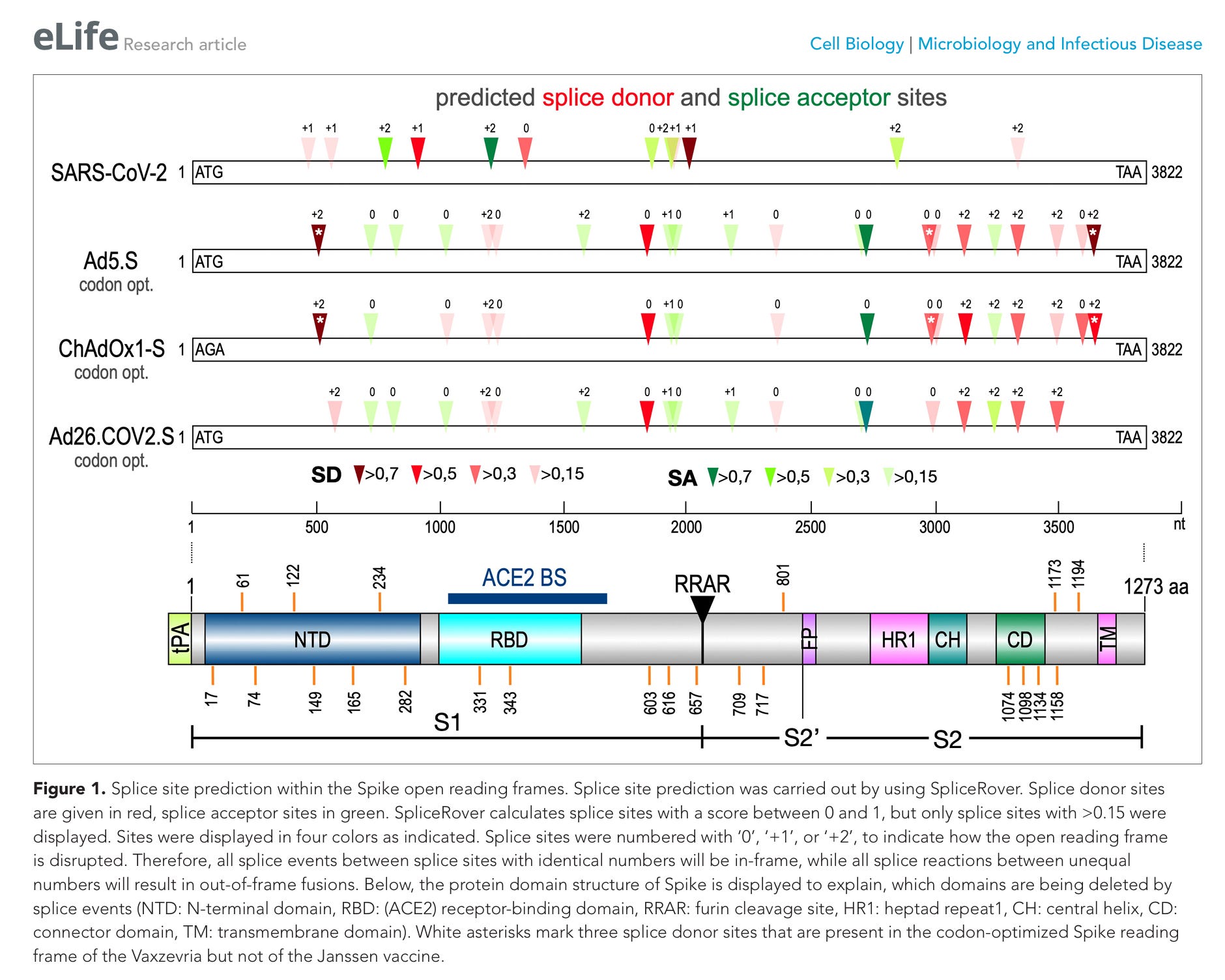
Abbildung 1. Vorhersage von Spleißstellen innerhalb der offenen Leserahmen des Spike. Die Vorhersage der Spleißstellen erfolgte unter Verwendung von SpliceRover. Spleiß-Donorstellen sind rot dargestellt, Spleiß-Akzeptorstellen grün. SpliceRover berechnet Spleißstellen mit einem Wert zwischen 0 und 1, es wurden jedoch nur Spleißstellen mit einem Wert > 0,15 angezeigt. Die Stellen wurden wie angegeben in vier Farben dargestellt. Die Spleißstellen wurden mit „0“, „+1“ oder „+2“ nummeriert, um anzugeben, wie der offene Leserahmen unterbrochen wird. Daher sind alle Spleißereignisse zwischen Spleißstellen mit identischen Nummern in-frame, während alle Spleißreaktionen zwischen ungleichen Nummern zu out-of-frame-Fusionen führen. Nachstehend wird die Proteindomänenstruktur von Spike dargestellt, um zu verdeutlichen, welche Domänen durch Spleißereignisse entfernt werden (NTD: N-terminale Domäne, RBD: (ACE2)-Rezeptorbindungsdomäne, RRAR: Furin-Spaltstelle, HR1: Heptad-Repeat 1, CH: zentrale Helix, CD: Verbindungsdomäne, TM: Transmembrandomäne). Weiße Sternchen markieren drei Spleiß-Donorstellen, die im codonoptimierten Spike-Leserahmen des Vaxzevria-Impfstoffs, nicht jedoch im Janssen-Impfstoff vorhanden sind.
Warum haben die Hersteller das Programm SpliceRover nicht über ihre Konstrukte laufen lassen? Wenn Pharmaunternehmen oder Forscher ein neues künstliches Gen (wie das Spike-Gen bei Johnson & Johnson oder AstraZeneca) designen, müssen sie verhindern, dass die menschliche Zelle dieses Gen fälschlicherweise alternativ spleißt. Werkzeuge wie SpliceRover (oder modernere Nachfolger wie SpliceAI) werden in der biomedizinischen Entwicklung genau dafür eingesetzt.
Erkennt das Tool ein hohes Spleiß-Risiko, hätten die Firmen die Sequenz entsprechend korrigieren müssen, um das zu unterbinden.
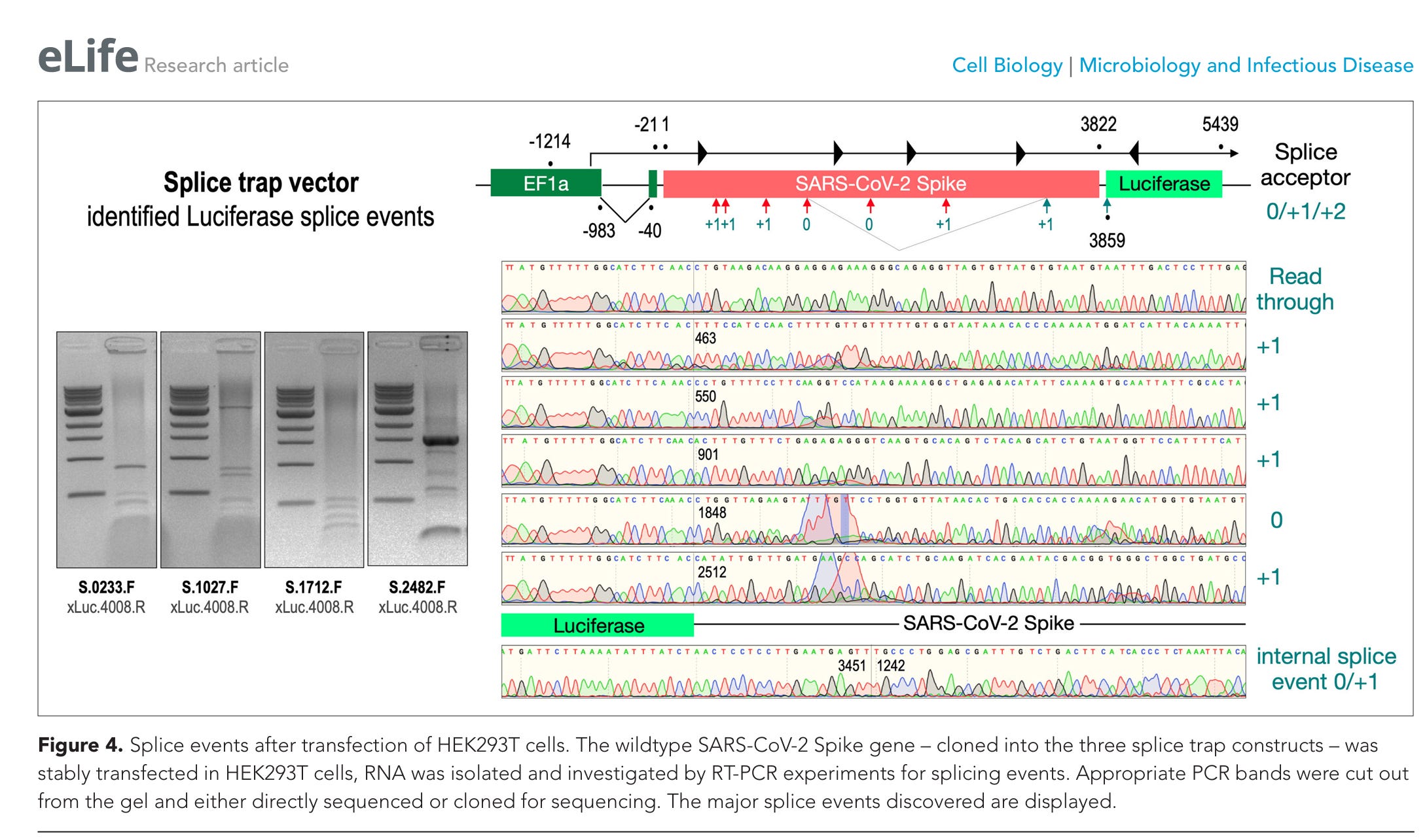
Abbildung 4. Spleißereignisse nach Transfektion von HEK293T-Zellen. Das Wildtyp-Spike-Gen von SARS-CoV-2 – das in die drei Spleißfalle-Konstrukte kloniert wurde – wurde stabil in HEK293T-Zellen transfiziert; anschließend wurde die RNA isoliert und mittels RT-PCR-Experimenten auf Spleißereignisse untersucht. Entsprechende PCR-Banden wurden aus dem Gel herausgeschnitten und entweder direkt sequenziert oder zur Sequenzierung kloniert. Die wichtigsten entdeckten Spleißereignisse sind dargestellt.
SPUTNIK V COMPONENT R-AD5-S
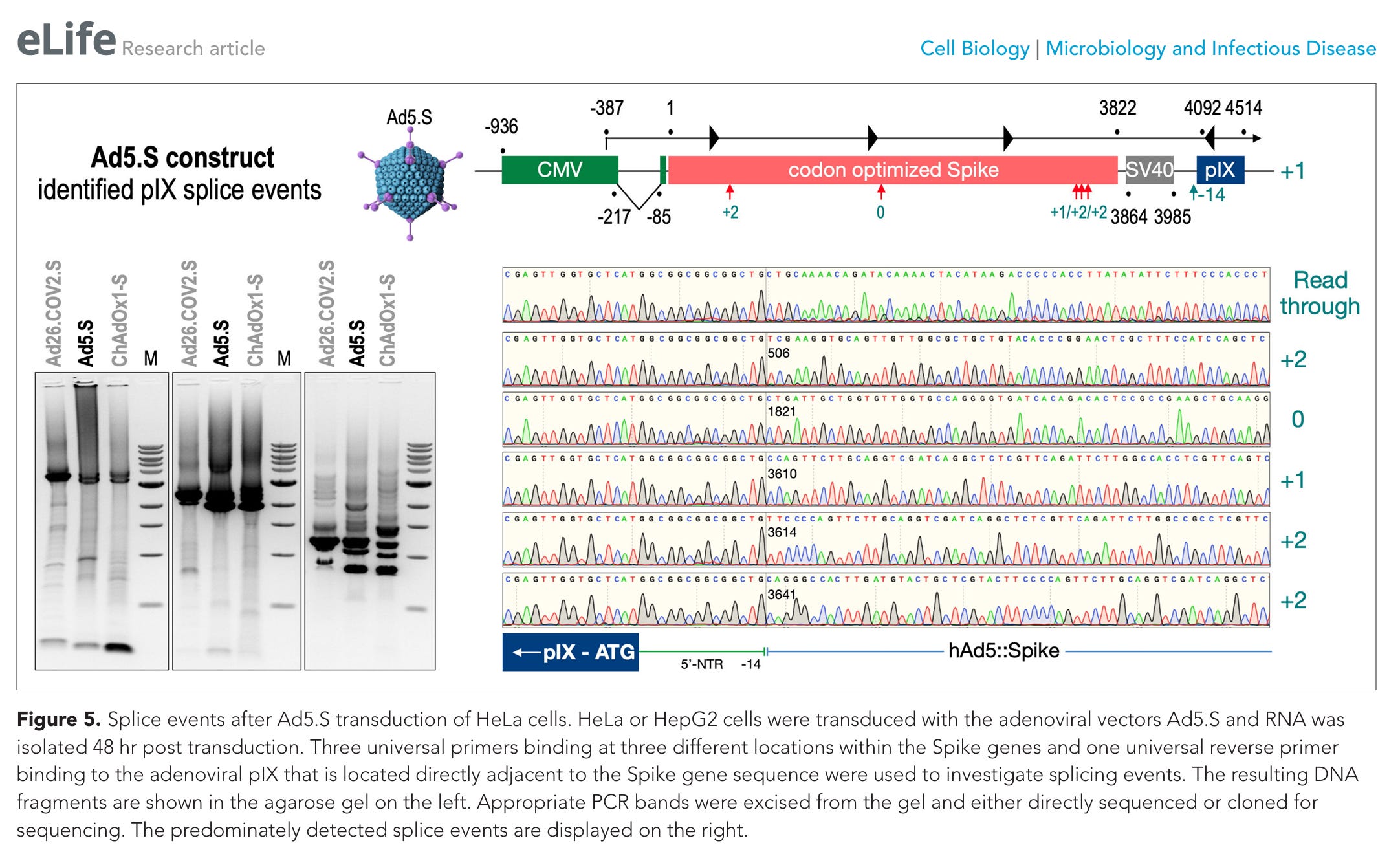
Abbildung 5. Spleißereignisse nach Ad5.S-Transduktion von HeLa-Zellen. HeLa- oder HepG2-Zellen wurden mit den adenoviralen Vektoren Ad5.S transduziert, und die RNA wurde 48 Stunden nach der Transduktion isoliert. Zur Untersuchung der Spleißereignisse wurden drei universelle Primer, die an drei verschiedenen Stellen innerhalb der Spike-Gene binden, sowie ein universeller reverser Primer verwendet, der an das adenovirale pIX bindet, das sich direkt neben der Spike-Gensequenz befindet. Die resultierenden DNA-Fragmente sind im Agarose-Gel auf der linken Seite dargestellt. Entsprechende PCR-Banden wurden aus dem Gel herausgeschnitten und entweder direkt sequenziert oder zur Sequenzierung kloniert. Die vorwiegend nachgewiesenen Spleißereignisse sind auf der rechten Seite dargestellt.
Vaxzevria, COVID-19 Vaccine (ChAdOx1-S [recombinant])
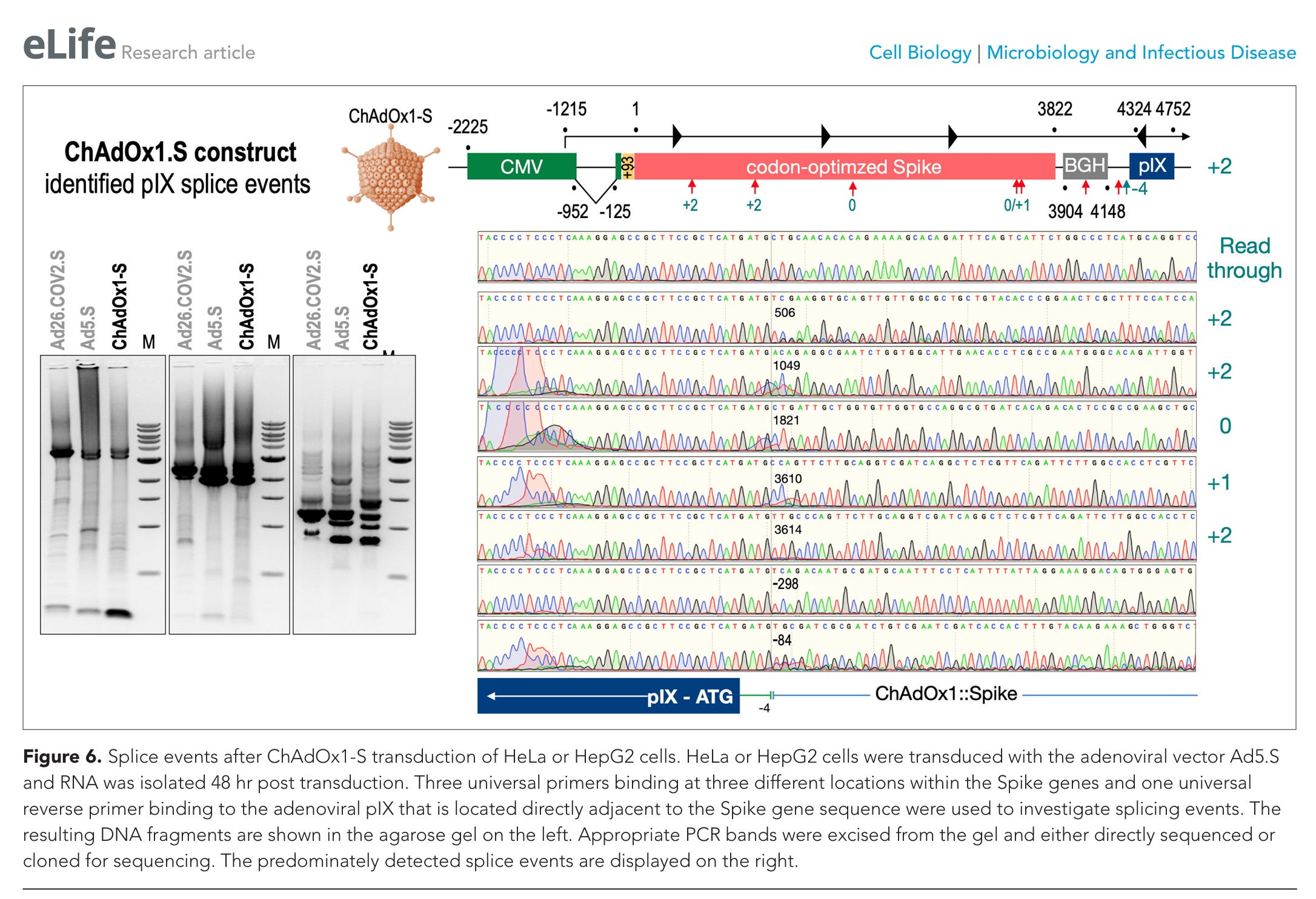
Abbildung 6. Spleißereignisse nach der Transduktion von HeLa- oder HepG2-Zellen mit ChAdOx1-S. HeLa- oder HepG2-Zellen wurden mit dem adenoviralen Vektor Ad5.S transduziert, und 48 Stunden nach der Transduktion wurde RNA isoliert. Zur Untersuchung der Spleißereignisse wurden drei universelle Primer, die an drei verschiedenen Stellen innerhalb der Spike-Gene binden, sowie ein universeller reverser Primer verwendet, der an das adenovirale pIX bindet, das sich direkt neben der Spike-Gensequenz befindet. Die resultierenden DNA-Fragmente sind im Agarose-Gel auf der linken Seite dargestellt. Entsprechende PCR-Banden wurden aus dem Gel herausgeschnitten und entweder direkt sequenziert oder zur Sequenzierung kloniert. Die vorwiegend nachgewiesenen Spleißereignisse sind auf der rechten Seite dargestellt.
Ad26.COV2-S=JCOVDEN®, vormals COVID-19 Vaccine Janssen
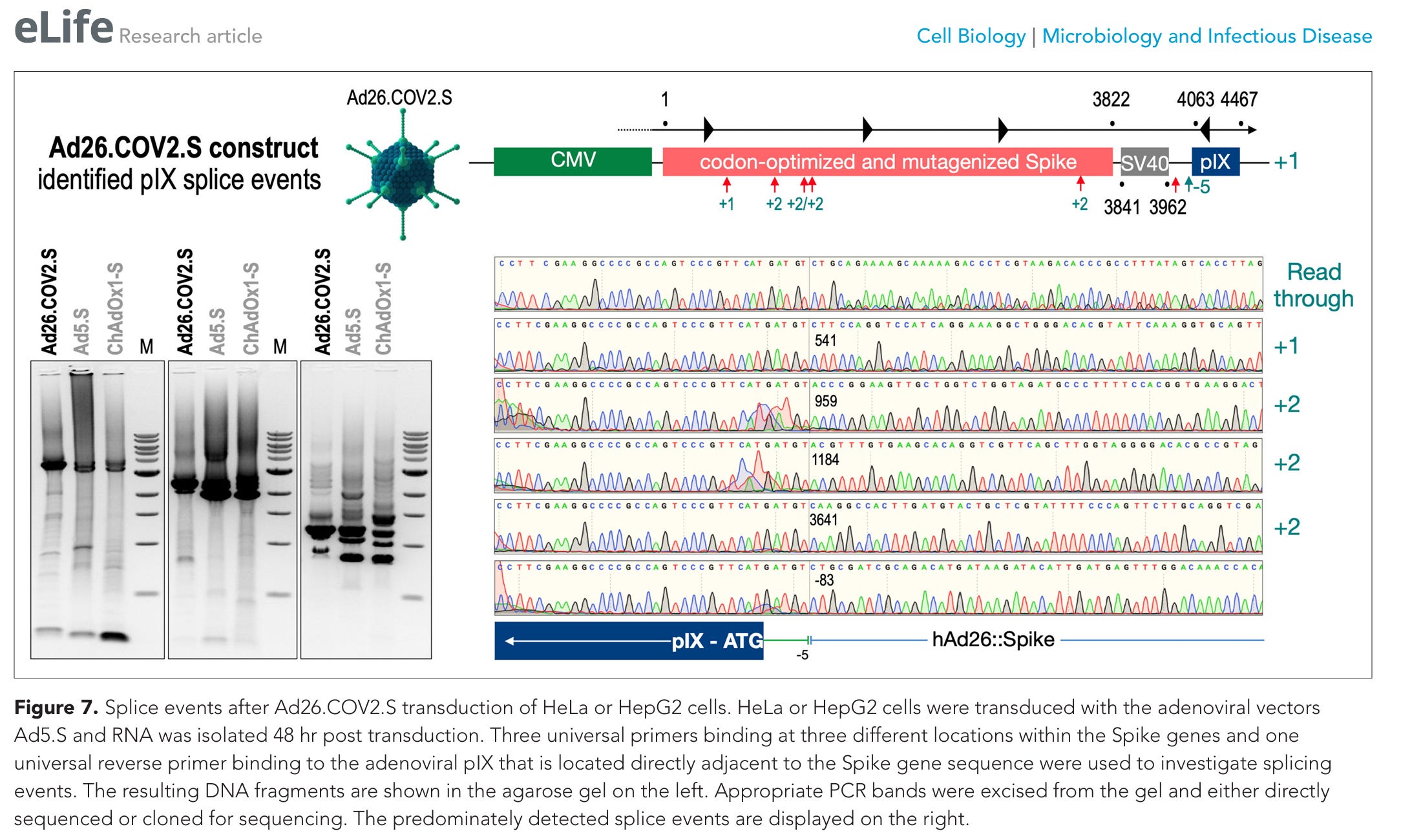
Abbildung 7. Spleißereignisse nach der Transduktion von HeLa- oder HepG2-Zellen mit Ad26.COV2.S. HeLa- oder HepG2-Zellen wurden mit den adenoviralen Vektoren Ad5.S transduziert, und die RNA wurde 48 Stunden nach der Transduktion isoliert. Zur Untersuchung der Spleißereignisse wurden drei universelle Primer, die an drei verschiedenen Stellen innerhalb der Spike-Gene binden, sowie ein universeller reverser Primer verwendet, der an das adenovirale pIX bindet, das sich direkt neben der Spike-Gensequenz befindet. Die resultierenden DNA-Fragmente sind im Agarose-Gel auf der linken Seite dargestellt. Entsprechende PCR-Banden wurden aus dem Gel herausgeschnitten und entweder direkt sequenziert oder zur Sequenzierung kloniert. Die vorwiegend nachgewiesenen Spleißereignisse sind auf der rechten Seite dargestellt.
Diskussion
Hier liefern wir experimentelle Belege dafür, dass Adenovirus-basierte Impfstoffe, die darauf ausgelegt sind, das SARS-CoV-2-Spike-Protein aus einer klonierten, codonoptimierten cDNA zu exprimieren, zu unerwünschten Spleißereignissen führen können, wenn bei der Vektorkonstruktion nicht darauf geachtet wird, dass keine funktionellen Spleißstellen in der kodierenden Sequenz vorhanden sind. […]
In dieser Studie haben wir unsere Hypothese experimentell validiert, dass Spike-Protein-Varianten, denen der natürliche Membrananker fehlt – und die dadurch der Sekretion unterliegen –, aus adenoviralen Impfstoffvektoren als Folge von RNA-Spleißereignissen im Zellkern entstehen. […]
Unsere Ergebnisse zeigen eindeutig, dass Spike-Spleißreaktionen stattfinden und dass sekretierte, lösliche Spike-Proteinvarianten entstehen. Western-Blot-Analysen von Gesamtzellen und deren Überstand ergaben eine Reihe von Spike-Proteinvarianten, die sich in ihrem Molekulargewicht unterscheiden, was auf die zuvor analysierten Spleißereignisse hindeutet. Es wurde beschrieben, dass lösliches Spike-Protein unerwünschte Wirkungen hervorruft, beispielsweise eine starke Entzündungsreaktion auf Endothelzellen (Lei et al., 2020; Nuovo et al., 2021; Patra et al., 2020; Amraei und Rahimi, 2020). Offensichtlich treten schwere, lebensbedrohliche thromboembolische Ereignisse aufgrund einer Infektion der Empfängerzellen durch Wildtyp-SARS-CoV-2 auf. Selbst Pseudoviren mit Spike-Protein an ihrer Oberfläche können starke Entzündungsreaktionen in Geweben und Endothelzellen auslösen, was auf ein potenzielles Risiko hinweist, dass lösliches Spike-Protein schwere Nebenwirkungen verursachen kann, wenn es systemisch im Gefäßsystem vorhanden ist (Lei et al., 2021).
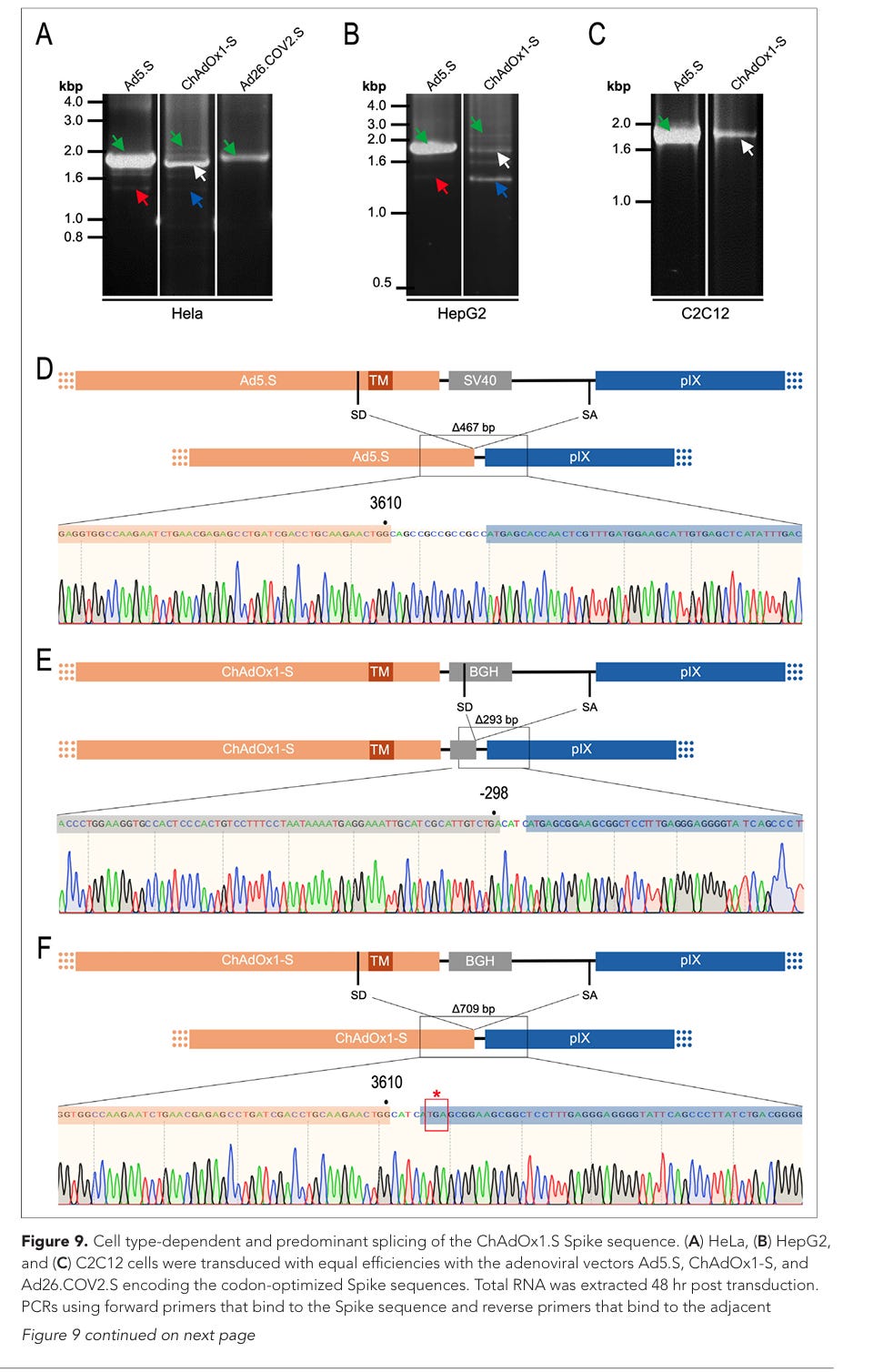
Abbildung 9. Zelltypabhängiges und vorherrschendes Spleißen der ChAdOx1.S-Spike-Sequenz. (A) HeLa-, (B) HepG2- und (C) C2C12-Zellen wurden mit gleicher Effizienz mit den adenoviralen Vektoren Ad5.S, ChAdOx1-S und Ad26.COV2.S transduziert, die für die codonoptimierten Spike-Sequenzen kodieren. 48 Stunden nach der Transduktion wurde die Gesamt-RNA extrahiert. Zur PCR-Amplifikation der erzeugten cDNAs wurden PCRs mit Vorwärtsprimern, die an die Spike-Sequenz binden, und Rückwärtsprimern, die an das angrenzende stromabwärts gelegene pIX im adenoviralen DNA-Genom binden, durchgeführt. Trennung der PCR-Amplikate mittels Agarosegel-Elektrophorese. Grüner Pfeil: Fragmente in voller Länge. Roter Pfeil: Gespaltenes Fragment der von Ad5.S kodierten Spike-Sequenz, das zu einem potenziell sekretierten Spike-Fusionsprotein führt. Weißer Pfeil: Gespaltenes Fragment der von ChAdOx1-S kodierten Spike-Sequenz, das zu einem Spike-pIX-Fusionsprotein führt. Blauer Pfeil: Gespaltenes Fragment der von ChAdOx1-S kodierten Spike-Sequenz, das zu einem potenziell sekretierten Spike-Protein führt. (D) Sequenzierungsergebnis des mit dem roten Pfeil markierten DNA-Fragments. (E) Sequenzierungsergebnis des mit dem weißen Pfeil markierten DNA-Fragments. (F) Sequenzierungsergebnis des mit dem blauen Pfeil markierten DNA-Fragments.
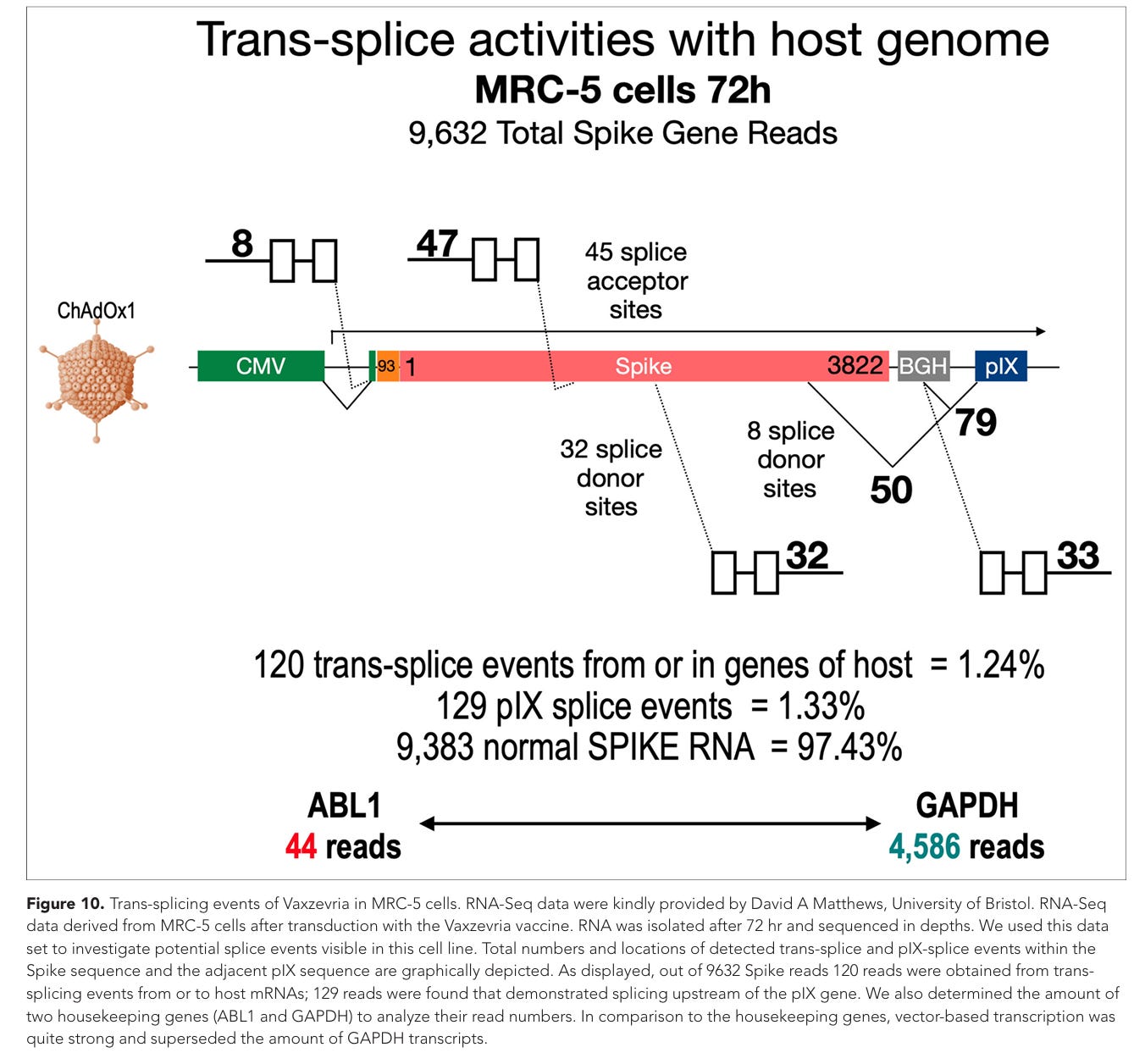
Abbildung 10. Trans-Spleißereignisse von Vaxzevria in MRC-5-Zellen. Die RNA-Seq-Daten wurden freundlicherweise von David A. Matthews von der Universität Bristol zur Verfügung gestellt. Die RNA-Seq-Daten stammen aus MRC-5-Zellen nach Transduktion mit dem Impfstoff Vaxzevria. Die RNA wurde nach 72 Stunden isoliert und mit hoher Sequenztiefe sequenziert. Wir nutzten diesen Datensatz, um potenzielle Spleißereignisse zu untersuchen, die in dieser Zelllinie sichtbar sind. Die Gesamtanzahl und die Positionen der nachgewiesenen Trans-Spleiß- und pIX-Spleißereignisse innerhalb der Spike-Sequenz und der angrenzenden pIX-Sequenz sind grafisch dargestellt. Wie dargestellt, stammten von 9632 Spike-Reads 120 Reads aus Trans-Spleißereignissen von oder zu Wirts-mRNAs; es wurden 129 Reads gefunden, die ein Spleißen stromaufwärts des pIX-Gens zeigten. Wir haben außerdem die Menge zweier Haushaltegene (ABL1 und GAPDH) bestimmt, um deren Read-Zahlen zu analysieren. Im Vergleich zu den Haushaltegene war die vektorbasierte Transkription recht stark und übertraf die Menge an GAPDH-Transkripten.
Dies wurde durch die Beobachtung weiter bestätigt, dass im Gegensatz zu Ad5.S und ChAdOx1-S bei der Zelltransduktion mit Ad26.COV2.S keine nennenswerten Mengen an gespleißten RNA-Molekülen entstanden. Dies könnte die im Vergleich zum Vaxzevria-Impfstoff etwa dreimal geringere Häufigkeit thromboembolischer Nebenwirkungen beim Janssen-Impfstoff erklären. Dies könnte auch das etwa zweifach geringere Risiko für TTS beim Janssen-Impfstoff erklären.
Es könnte aber auch einfach daran liegen, dass der AstraZeneca “Impfstoff” massiv mit Zellkultur verunreinigt war. Das wusste man zum Publikationszeitpunkt bereits, ignoriert es jedoch leider. Zudem gibt es noch ein weiteres Problem mit dem Core Protein VII. Das Problem mit Core Protein VII war J&J 2022 bereits bekannt, aber nicht öffentlich bekannt.
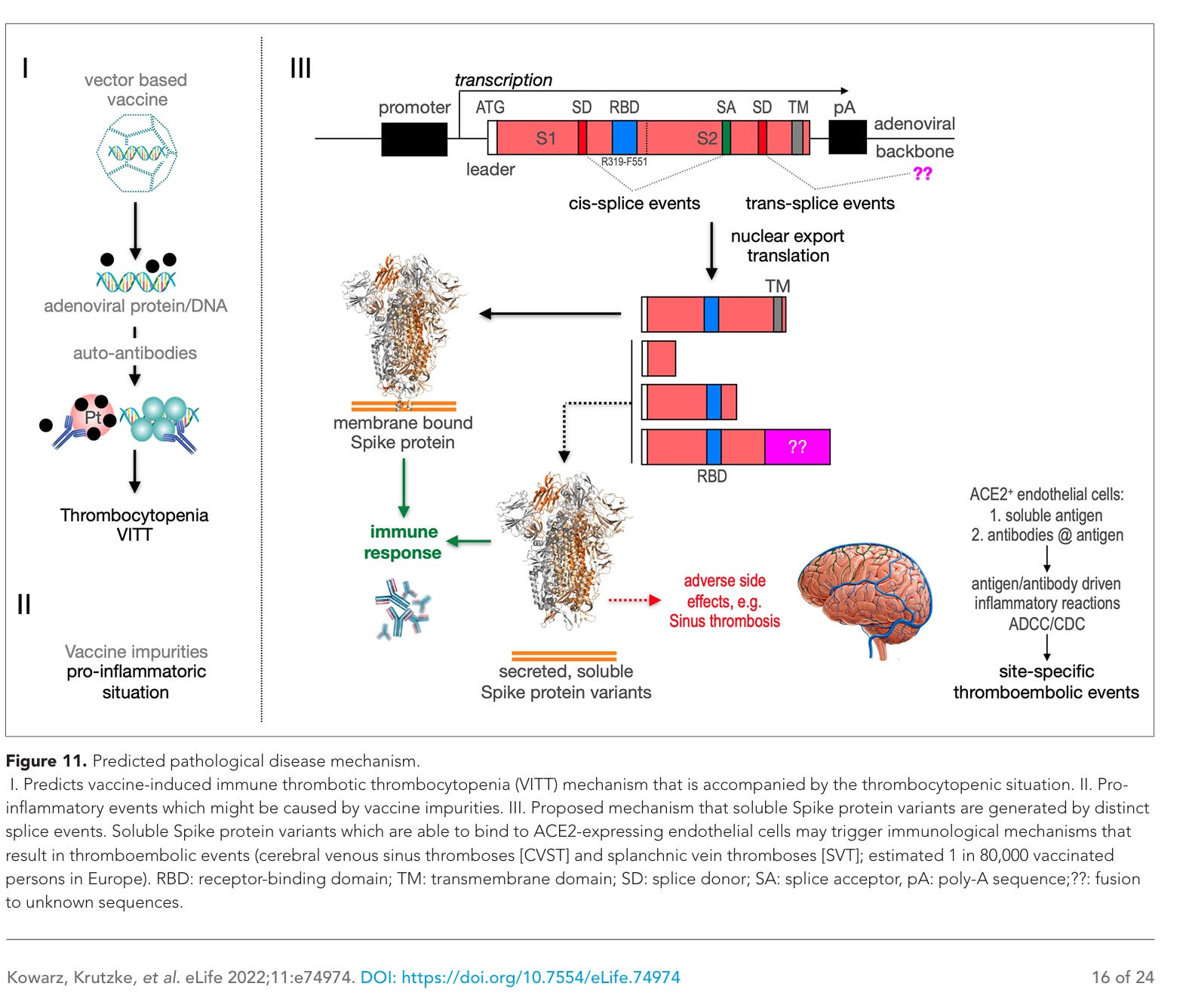
Abbildung 11. Vorgestellter pathologischer Krankheitsmechanismus.
I. Vorgestellter Mechanismus der impfstoffinduzierten immunologischen thrombotischen Thrombozytopenie (VITT), die mit einer Thrombozytopenie einhergeht.
II. Proinflammatorische Ereignisse, die möglicherweise durch Verunreinigungen im Impfstoff verursacht werden.
III. Vorgeschlagener Mechanismus, wonach lösliche Spike-Protein-Varianten durch unterschiedliche Spleißereignisse entstehen. Lösliche Spike-Protein-Varianten, die an ACE2-exprimierende Endothelzellen binden können, könnten immunologische Mechanismen auslösen, die zu thromboembolischen Ereignissen führen (zerebrale Venensinusthrombosen [CVST] und splanchnische Venenthrombosen [SVT]; geschätzt 1 von 80.000 geimpften Personen in Europa). RBD: Rezeptorbindungsdomäne; TM: Transmembrandomäne; SD: Spleißdonor; SA: Spleißakzeptor; pA: Poly-A-Sequenz; ??: Fusion mit unbekannten Sequenzen
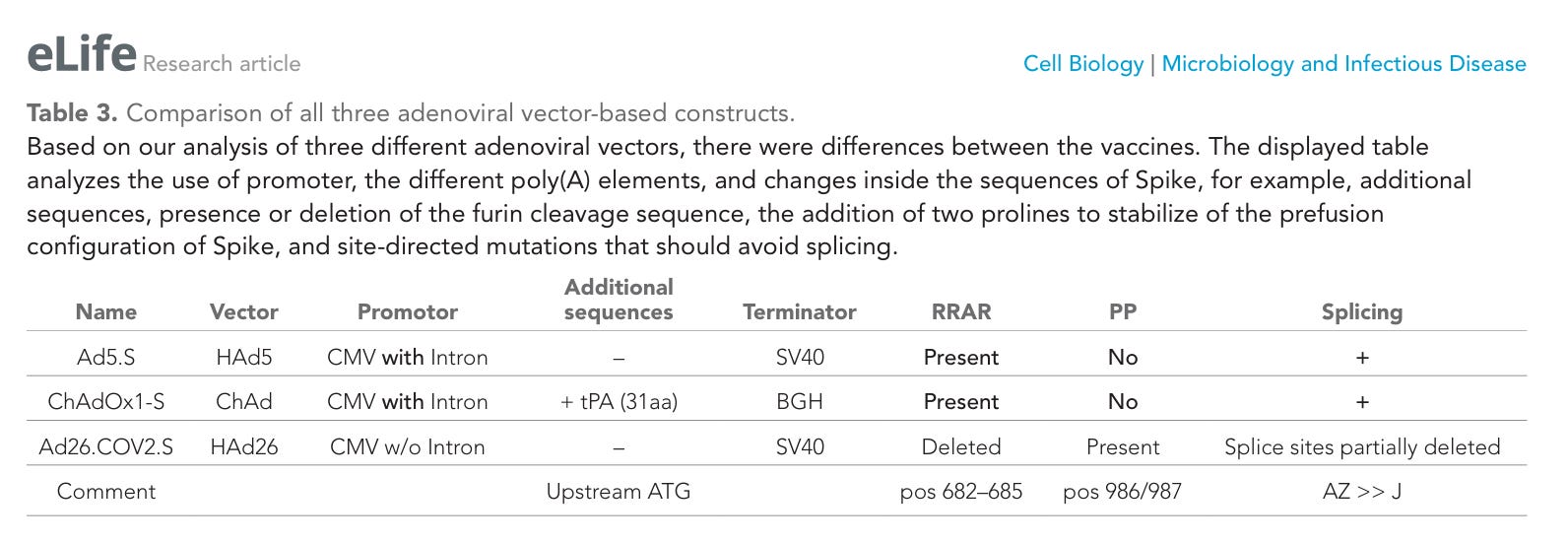
Tabelle 3. Vergleich aller drei auf adenoviralen Vektoren basierenden Konstrukte. Unsere Analyse der drei verschiedenen adenoviralen Vektoren ergab Unterschiede zwischen den Impfstoffen. Die dargestellte Tabelle analysiert die Verwendung des Promotors, die verschiedenen Poly(A)-Elemente und Veränderungen innerhalb der Spike-Sequenzen, beispielsweise zusätzliche Sequenzen, das Vorhandensein oder die Deletion der Furin-Spaltsequenz, die Hinzufügung von zwei Prolinen zur Stabilisierung der Präfusionskonfiguration von Spike sowie ortsspezifische Mutationen, die ein Spleißen verhindern sollen.
Über das Verbleib und die Gewebeverteilung von Adenovirus-basierten Impfstoffen beim Menschen ist nur sehr wenig bekannt. Bei Mäusen wurde beobachtet, dass nach intramuskulärer Injektion erhebliche Mengen an Adenovirus-Vektoren in die Hepatozyten gelangen, wo die Expression des Transgens stattfindet (Kron et al., 2011). Es ist wahrscheinlich, dass das Schimpansen-ChAdOx1-S ebenfalls ein hepatotropes Adenovirus ist, da es auf dem Schimpansen-Adenovirus Y25 basiert (Dicks et al., 2012), das zu einer Gruppe von Isolaten aus Schimpansen gehörte, die mit Ausbrüchen viraler Hepatitis in Verbindung standen (Hillis und Goodman, 1969). Sollte es in (einigen der) Impfstoffe nach intramuskulärer Injektion in nennenswertem Umfang zu einer Transduktion von Hepatozyten kommen, ist es denkbar, dass größere Mengen an Spike-Proteinen, die durch Spleißen die Transmembrandomäne verloren haben, über die Lebervenen in den systemischen Kreislauf ausgeschüttet werden.
Zusammenfassend lässt sich aus unserer Arbeit ableiten, dass bei DNA-kodierten Sequenzen (insbesondere wenn sie von RNA-Viren stammen) vor allem starke SDs vermieden und starke Poly(A)-Signale für die RNA-Termination in Expressionskonstrukten verwendet werden sollten. Dies ist besonders wichtig, wenn adenovirale Vektoren als Transportvehikel eingesetzt werden. In der Regel werden Transgensequenzen in das adenovirale Genom in die Lücke der deletierten E1-Region eingebettet, die sich direkt stromaufwärts des adenoviralen pIX mit einer starken SA-Stelle befindet. Alternativ würde, während die meisten Transgene in adenoviralen Vektoren eine „Links-nach-Rechts“-Orientierung aufweisen, auch eine Inversion der Expressionskassette Spleißereignisse zwischen der transgenen kodierenden Sequenz und dem viralen pIX-Transkript verhindern. Unter Berücksichtigung dieser Tatsache wird die Wahrscheinlichkeit eines Spleißens stark reduziert oder auf Null gesenkt. Wir betonen nachdrücklich die Berücksichtigung dieser Aspekte für die rationale Entwicklung sicherer adenoviraler Impfstoffvektoren in der Zukunft sowie für adenovirale vektorbasierte Gentransferansätze im Allgemeinen.
Wenn du einen Bauplan nimmst, der eigentlich für eine Holzkonstruktion gedacht war (ein RNA-Virus), und diesen jetzt eins zu eins für ein Stahlgebäude (DNA) nutzt, gibt das Probleme. Wenn du dann noch billige Verbindungsstücke (schwache Poly(A)-Signale / Stoppschilder) verbaust, hält die Bremse nicht. Die Maschine läuft über das Ziel hinaus und vermischt sich mit dem Nachbarbauteil (dem viralen pIX).
Damit in Zukunft kein Pfusch am Bau entsteht, müssen wir drei Dinge ändern:
Regel 1: Keine falschen Kanten einbauen (Starke SDs vermeiden). Wir müssen verhindern, dass das Material „Schnittstellen“ (SDs) hat, an denen die Zelle fälschlicherweise anfängt zu sägen und neu zusammenzusetzen.
Regel 2: Eine ordentliche Bremse einbauen (Starke Poly(A)-Signale nutzen). Wir brauchen am Ende des Bauteils ein fettes, stabiles Stoppschild. Das sorgt dafür, dass die Maschine exakt dort aufhört zu arbeiten und nicht in die benachbarten Bauteile hineinrast.
Regel 3: Das Bauteil umdrehen (Inversion der Expressionskassette). Normalerweise bauen wir Teile von links nach rechts ein. Wenn wir dieses spezifische Gen-Bauteil aber einfach spiegelverkehrt (von rechts nach links) einsetzen, können die Funken des einen Teils das Nachbarteil biologisch überhaupt nicht mehr erreichen. Das Risiko für Fehlkonstruktionen (Spleißen) sinkt damit sofort auf Null.
Das Fazit des Meisters:
„Wir haben gelernt, wie man diese Gen-Taxis richtig konstruiert. Wenn sich in Zukunft alle Entwickler an diese drei Bauregeln halten, sind Fehler beim Zusammenbau (Spleißen) komplett ausgeschlossen. Das gilt für neue Impfstoffe genauso wie für jede andere Gen-Therapie.“
Im Klartext, alle Hersteller haben bei der Konstruktion Mist gebaut und Schulstoff ignoriert, aber bis heute wird das Problem ignoriert und keiner redet darüber.
Unterstützungsmöglichkeiten:
Bücherwunschzettel: https://www.amazon.de/registries/gl/owner-view/30LG3DJ4ET90L?ref_=list_d_gl_lfu_nav
Andere Unterstützungsmöglichkeiten für Holgers und meine Forschung:
Konto für Unterstützung für das Projekt Scan 2000
Dr. Merse DE34 4305 0001 0302 7851 75 Sparkasse Bochum
Horst Reissner: IBAN DE51 4401 0046 0406 4514 67
Dr. S. Stebel: https://ko-fi.com/einmalmitprofisarbeiten
Paypal: [email protected]
Wikipedia-Autoren. (2003, April 7). Alternatives spleißen. https://de.wikipedia.org/wiki/Alternatives_Splei%C3%9Fen
DocCheck, M. B. (n.d.). Alternatives spleißen – DocCheck Flexikon. DocCheck Flexikon. https://flexikon.doccheck.com/de/Alternatives_Splei%C3%9Fen
Wikipedia-Autoren. (2003, April 7). Alternatives spleißen. https://de.wikipedia.org/wiki/Alternatives_Splei%C3%9Fen
User, S. (2024, April 23). Genetik: splicing. Abitur-Wissen.org. https://abitur-wissen.org/index.php/biologie/genetik/821-genetik-splicing

Zuallaert, J., Godin, F., Kim, M., Soete, A., Saeys, Y., & De Neve, W. (2018). SpliceRover: interpretable convolutional neural networks for improved splice site prediction. Bioinformatics, 34(24), 4180–4188. https://doi.org/10.1093/bioinformatics/bty497
Zabardast, A., Tamer, E. G., Son, Y. A., & Yılmaz, A. (2023). An automated framework for evaluation of deep learning models for splice site predictions. Scientific Reports, 13(1), 10221. https://doi.org/10.1038/s41598-023-34795-4
R-AD5-S (RECOMBINANT ADENOVIRUS TYPE 5 CARRYING THE GENE FOR SEVERE ACUTE RESPIRATORY SYNDROME CORONAVIRUS 2 (SARS-COV-2) SPIKE GLYCOPROTEIN) https://precision.fda.gov/ginas/app/ui/substances/6cf5c858-21a4-46e8-88d9-40624fac1d51
Greinacher, A., Selleng, K., Palankar, R., Wesche, J., Handtke, S., Wolff, M., Aurich, K., Lalk, M., Methling, K., Völker, U., Hentschker, C., Michalik, S., Steil, L., Reder, A., Schönborn, L., Beer, M., Franzke, K., Büttner, A., Fehse, B., . . . Renné, T. (2021). Insights in ChAdOx1 nCoV-19 vaccine-induced immune thrombotic thrombocytopenia. Blood, 138(22), 2256–2268. https://doi.org/10.1182/blood.2021013231
Krutzke, L., Rösler, R., Allmendinger, E., Engler, T., Wiese, S., & Kochanek, S. (2022). Process- and product-related impurities in the ChAdOx1 nCov-19 vaccine. eLife, 11. https://doi.org/10.7554/elife.78513
Wang JJ, Schönborn L, Warkentin TE, Müller L, Thiele T, Ulm L, Völker U, Ameling S, Franzenburg S, Kaderali L, Tzvetkova A, Colella A, Chataway T, Tan CW, Armour B, Troelnikov A, Rutten L, McCluskey J, Zahn R, Gordon TP, Greinacher A. Adenoviral Inciting Antigen and Somatic Hypermutation in VITT. N Engl J Med. 2026 Feb 12;394(7):669-683. doi: 10.1056/NEJMoa2514824. PMID: 41671482; PMCID: PMC12900036. https://pubmed.ncbi.nlm.nih.gov/41671482/